3. Base química de la herencia.
3. Base química de la herencia.
La genética molecular se ocupa de la naturaleza de los genes y de su expresión. Los genes están compuestos por moléculas de ADN que se replican antes de la mitosis. Cada una de las copias originadas en esta replicación contiene la información necesaria para el desarrollo del nuevo ser; por tanto, lo que se hereda es fundamentalmente ADN, una molécula informativa. Las funciones del ADN son la conservación y la transmisión de la información, funciones ambas que requieren energía ya que, desde el punto de vista termodinámico, la información es orden, es decir, entropía negativa. La expresión de los genes, el fenotipo, es, en última instancia, una o varias proteínas que desempeñan unas funciones específicas, las cuales dependen de su forma, de su conformación. Esta se halla determinada por la secuencia de aminoácidos que la componen y la secuencia de aminoácidos está determinada a su vez por la secuencia de nucleótidos del ADN. Los genes son fragmentos de ADN que se expresan cuando la información que contienen se traduce para formar proteínas con una función biológica específica.
El descubrimiento de que el ADN era la molécula portadora de la información ge nética ha sido la revolución de la biología del siglo xx.
3.1. El ADN como portador de la información genética.
Hoy sabemos que la molécula que contiene la información de las características biológicas de los seres vivos es el ADN. Sin embargo, la demostración de que este ácido nucleico constituía el material hereditario sólo fue posible gracias a la paciente labor de investigación de muchos científicos durante la primera mitad del siglo xx.
Antes de que se identificara la molécula portadora del mensaje genético, ya se sabía que ésta debía cumplir ciertos requisitos:
- Tenía que ser químicamente estable para que la información contenida en la molécula no sufriera alteraciones.
- Debía ser capaz de replicarse y originar copias de sí misma que pasaran a las células hijas durante la división celular, asegurando de esta manera la pervivencia de la información biológica de una célula determinada en su estirpe.
- Era necesario, además que esa información pudiera transmitirse de una generación a otra para permitir que las características biológicas pasaran a la descendencia.
- Por último, aunque fuera químicamente estable, la molécula debía ser susceptible de sufrir cambios que posibilitaran la aparición de cierta variabilidad a fin de poder explicar la evolución de los seres vivos.
Aunque el ADN ya se conocía desde su descubrimiento en 1869 por el científico suizo Friedrich Miescher, se consideraba que eran las proteínas, y no el ADN, las portadoras de la información genética.
No obstante, el descubrimiento de que los cromosomas se dividían y transmitían durante la división celular en las células eucariotas, permitió comprobar que ambos componentes cromosómicos (ADN y proteínas) cumplían los requisitos citados.
Ya en 1928, en el curso de sus experimentos con la bacteria Streptococcus pneumoniae, agente causal de la neumonía, Frederick Griffith demostró que la capacidad biológica de producir una cápsula (hecho que determina la virulencia de las bacterias) podía ser adquirida de otra cepa por medio de una sustancia, aún no identificada, a la que se denominó «factor transformante».
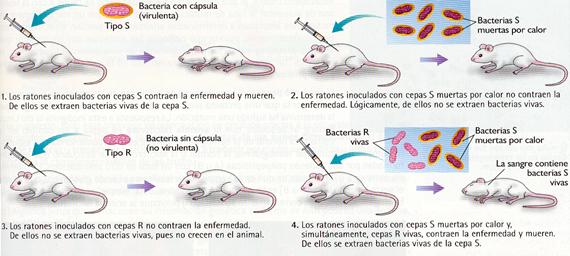
Experimentos de Griffith sobre la transformación bacteriana.
En 1944, Oswald T. Avery, Maclyn McCarty y Colin MacLeod observaron que la capacidad transformante de las cepas virulentas de Streptococcus pneumoniae desaparecía cuando se agregaban enzimas que destruían el ADN. Dedujeron de este hecho que el factor transformante era la molécula de ADN.
Existían, además, otros indicios que apoyaban esta idea. Por una parte, se había comprobado que la cantidad de ADN era la misma para todas las células somáticas de los individuos de una determinada especie, mientras que los gametos sólo tenían la mitad. Así mismo, los estudios de Erwin Chargaff sobre las similitudes en las proporciones de bases nitrogenadas presentes en el ADN de los individuos de la misma especie parecían confirmar la relación existente entre esta molécula y la información genética.
La prueba definitiva fue obtenida en 1952 por Alfred Hershey y Martha Chase, quienes demostraron de forma concluyente que el ADN, y no una proteína, era el material genético del bacteriófago T2. Al año siguiente, James Watson y Francis Crick elaboraron su famoso modelo de doble hélice para explicar la estructura de la molécula del ADN.
3.1.1. ADN y cromosomas.
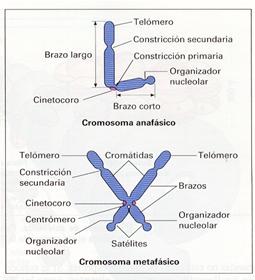
Los cromosomas son estructuras cilíndricas que representan el grado más elevado de empaquetamiento del ADN y, por tanto, de la cromatina en la célula. Durante la metafase, cada cromosoma está formado por dos estructuras idénticas, llamadas cromátidas, que confieren al cromosoma la característica apariencia simétrica en X. Las cromátidas están unidas entre sí por una región denominada centrómero o constricción primaria, llamada así porque aparece como un estrechamiento (constricción) del cromosoma. En el centrómero se encuentra el cinetocoro, que es la región del cromosoma por la que éste se une al huso mitótico. Las partes del cromosoma a cada lado del centrómero se denominan brazos. Según la posición del centrómero los cromosomas se clasifican en:
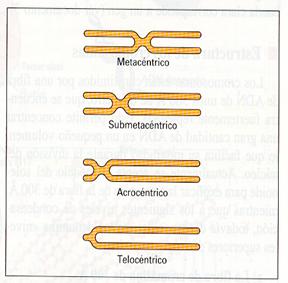
- Telocéntricos: el centrómero se sitúa en un extremo del cromosoma, de manera que puede decirse que éste posee un único brazo.
- Subtelocéntrico o acrocéntrico: uno de los brazos es mucho más corto que el otro.
- Submetacéntrico: hay cierta diferencia, aunque no mucha, de longitud entre los brazos.
- Metacéntrico: los brazos son prácticamente iguales.
Además del centrómero, se observan otros estrechamientos que son denominados constricciones secundarias. Se sabe que alguna de ellas constituye el organizado nucleolar.
El segmento final de cada cromátida es denominado telómero. Los telómeros parecen estar relacionadas con la estabilidad de los cromosomas. En ocasiones, un segmento final de la cromátida puede aparecer casi separado del resto por una constricción secundaria, recibiendo el nombre de satélite, o simplemente sat.
|
|
|
|
Morfología de los cromosomas anafásico y metafísico |
Tipos de cromosomas según la posicióndel centrómero |
3.1.2. Concepto de gen.
Un gen es un segmento de ADN que contiene la información necesaria para que, al ser transcrito y traducido, se sintetice una cadena polipeptídica.
A principios de siglo Garrod descubrió que ciertas enfermedades (como la alcaptonuria) estaban producidas por defectos metabólicos. Además, eran heredables de padres a hijos. En 1948 Beadle y Tatum emitieron la teoría de "un gen ---- un enzima ". Dedujeron que al fallar un gen (y faltar, por tanto un enzima) la ruta metabólica queda bloqueada en aquella sustancia sobre la que el enzima debería actuar.
Teoría un gen --- una enzima.
Debido a que hay enzimas formados por más de una cadena polipeptídica se prefiere formular la teoría "un gen ---- un polipéptido". Actualmente se sabe que los genes pueden ser discontinuos y constar de fragmentos de información dispersos a lo largo de la molécula de ADN (exones e intrones).
Gregor Mendel ya hablaba de “factores hereditarios” que se transmiten de generación en generación sin mezclarse ni alterarse y daban lugar a los caracteres de los individuos (genes). A principios de siglo, el conocimiento de los cromosomas y de los mecanismos de la mitosis y de la meiosis permitió deducir a Morgan que los genes se alojaban en los cromosomas. Éstos se transmiten íntegros de generación en generación, y cada individuo recibe un juego de cromosomas del padre y otro de la madre, que al transmitirlos a la descendencia vuelven a separarse y pasan a individuos distintos. Los individuos diploides tienen un par de genes para regir cada carácter, uno aportado por el padre y otro aportado por la madre. El carácter que poseerá el individuo dependerá de la interacción entre ambos genes, que ahora se llaman alelos.
Puede ocurrir que un alelo se exprese y otro no (dominante y recesivo: Herencia dominante) o que el carácter expresado sea intermedio al expresado por cada alelo (herencia intermedia).
A veces un carácter está codificado por más de una pareja de genes: alelismo múltiple. Otros genes, en homocigosis, producen la muerte del individuo que los posee, normalmente en estado embrionario: genes letales. Algunos caracteres son el resultado de la interacción entre dos o más genes, que pueden cooperar o modificar mutuamente su acción: interacción génica o epistasis .
3.1.3. Conservación de la información: replicación del ADN.
El ADN portador de la información genética debe transmitirse fielmente a cada una de las células hijas obtenidas tras la división celular. Por tanto, antes de producirse ésta, es imprescindible que el ADN pueda formar réplicas exactas de sí mismo para disponer de dos copias iguales. Este proceso, conocido como replicación o autoduplicación, resulta fundamental para asegurar que todas las células de un organismo pluricelular mantienen la misma identidad.
Cuando Watson y Crick elaboraron su modelo de doble hélice en 1953, indicaron también cuál podría ser el mecanismo para llevar a cabo la replicación del ADN: separación de las dos cadenas y síntesis de la cadena complementaria de cada una de ellas. Sin embargo, otros investigadores plantearon distintas hipótesis que dieron lugar a tres posibles formas de replicación:
- Conservativa . La doble cadena original se mantiene y se sintetiza otra completamente nueva.
- Semiconservativa . Es la propuesta por Watson y Crick. Una de las hebras de cada doble hélice procede de la original, mientras que la otra se sintetiza nuevamente.
- Dispersiva . En cada doble hélice existen fragmentos de la original y fragmentos nuevos.
Poco después, Matthew Meselson y Franklin Stahl demostraron experimentalmente que la hipótesis correcta era la semiconservativa.
Herbert Taylor confirmó, así mismo, esta hipótesis en células eucariotas y en 1963 J. Cairns visualizó el proceso en Escherichia coli con técnicas autorradiográficas.
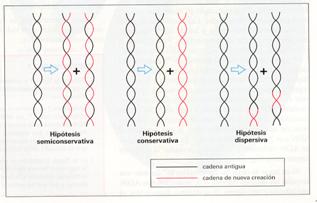
Hipótesis sobre la duplicación de ADN.
MECANISMO DE LA REPLICACIÓN
El mecanismo de la replicación es un proceso que ocurre una sola vez en cada generación celular, durante la fase S del ciclo celular.
El principio de la replicación según el cual cada cadena de la doble hélice de ADN sirve como molde para la formación de una nueva cadena, es relativamente simple; sin embargo, el proceso real es considerablemente complejo. Para su estudio se pueden diferenciar las siguientes etapas:
Inicio de la replicación
- La iniciación siempre comienza con una secuencia específica de nucleótidos conocida como origen de la replicación; requiere enzimas llamadas helicasas, las cuales rompen los enlaces de hidrógeno que mantienen unidas las bases complementarias, abriendo así la doble hélice.
- La separación de las cadenas, provoca superenrollamientos en las zonas vecinas, por lo que existen otras enzimas, las topoisomerasas o girasas que rebajan la tensión.
- Una vez separadas las dos cadenas, unas proteínas de unión a cadena simple (proteínas ssb) se unen a las hebras individuales, manteniéndolas separadas y evitando que se retuerzan.
Formación de las nuevas hebras
- Ahora bien, para que se forme una nueva cadena no es suficiente que esté presente la cadena vieja que sirve de molde, sino que debe estar presente el inicio de la nueva cadena; este inicio lo proporciona un fragmento de ARN llamado cebador (sintetizado por una ARN primasa, acoplado mediante una ARN-polimerasa y retirado por una ADN polimerasa), los cuales son reconocidos por las ADN polimerasas que se ponen a sintetizar la nueva cadena de ADN.
La síntesis real de las nuevas cadenas es catalizada por un grupo de enzimas conocidas como ADN polimerasas, que van añadiendo los nucleótidos uno a uno. La zona donde ocurre la replicación se observa al microscopio electrónico como un ojo o burbuja de replicación; estos segmentos de ADN en replicación se denominan replicones. En los extremos de la burbuja las cadenas forman una estructura en «Y» conocida como horquilla de replicación.
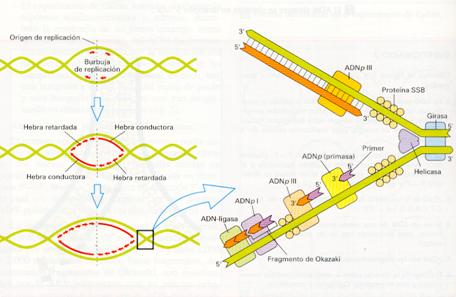
Mecanismos de duplicación del ADN.
- El proceso de replicación es bidireccional y siempre en el sentido de la cadena de nucleótidos 5'®3', pues las polimerasas sólo colocan y unen nucleótidos en ese sentido.
Como la replicación sólo ocurre en un sentido y las dos cadenas del ADN son antiparalelas, se planteaba un problema sobre cómo se efectuaría la replicación en los dos brazos de la horquilla. La solución la halló Reiji Okazaki, al encontrar que una cadena (la 5'--3') se sintetiza continuamente como una sola unidad, es la cadena adelantada o conductora, mientras que la otra cadena (la 3'--5') se forma de manera discontinua, como una serie de fragmentos de Okazaki, sintetizados cada uno en el sentido 5--3', que después terminan uniéndose formandose la llamada cadena retrasada o retardada.
- Por último, hay otra enzima, la ADN ligasa que conecta los fragmentos de ADN recién formados con la cadena de ADN en crecimiento.
CORRECCIÓN DE ERRORES
El ADN es la única molécula capaz de efectuar una reparación de sí misma. La replicación no ha concluido hasta que se comprueba que la copia de la secuencia nucleotídica es correcta. Es necesario, pues, detectar y corregir los errores producidos.
Aunque la ADN polimerasa III no une los nucleótidos que no sean complementarios de los correspondientes nucleótidos de la hebra molde, si se produce un error, el nucleótido mal emparejado es eliminado por la acción de enzimas exonucleasas . Por esta razón, el número de errores producidos durante el proceso de replicación es muy bajo (uno por cada 107-108 bases incorporadas).Sin embargo, esta proporción tan baja no es despreciable, ya que el número de nucleótidos de una cadena de ADN es muy alto, sobre todo en organismos pluricelulares (con más información genética y un gran número de células). Por ello, existe un proceso posreplicativo de corrección de errores en el que participan varias enzimas:
- Endonucleasas que detectan errores y cortan la cadena anómala.
- Exonucleasas que eliminan el fragmento incorrecto.
- ADN polimerasas que sintetizan la parte correspondiente al segmento eliminado. Esta acción y la anterior pueden ser realizadas por la ADN polimerasa I, como ya se indicó.
- ADN ligasas que unen el nuevo segmento al resto de la cadena.
|
|
|
|
|
|
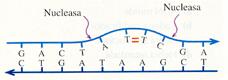
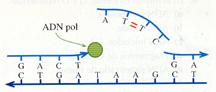
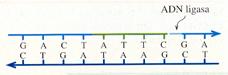
Eliminación de errores.
Tras la corrección, el número de errores desciende hasta uno por cada 1010 bases incorporadas.
Para posibilitar la detección de los errores es necesario diferenciar la cadena nueva de la antigua. Esto se consigue por metilación de las adeninas, proceso que tiene lugar pasado un cierto tiempo. Así, las adeninas pertenecientes a la hebra recién sintetizada no están aún metiladas y las de la hebra antigua sí, lo que permite que las enzimas reparadoras la identifiquen.
A pesar de los mecanismos correctores de errores, la fidelidad en la replicación no es absoluta, lo cual no es necesariamente negativo, ya que si los errores no tienen consecuencias sobre la viabilidad de las células (o de los individuos) que los poseen, se convierten en fuente de variación genética, imprescindible para el desarrollo de los procesos evolutivos.
Así, aunque en la replicación resulta fundamental mantener la fidelidad del mensaje genético en la síntesis de nuevas copias de ADN, se deja siempre un pequeñísimo margen a la aparición de variaciones que contribuyen a los cambios evolutivos.
DIFERENCIAS ENTRE EL PROCESO REPLICATIVO EN PROCARIOTAS Y EUCARIOTAS
Las diferencias en la replicación del ADN entre las células procariotas y eucariotas no afectan al mecanismo fundamental. Entre estas diferencias se pueden citar las siguientes:
- Como el ADN de los eucariotas está asociado con las histonas, la replicación debe tener en cuenta la síntesis de estas proteínas.
- El tamaño de los fragmentos de Okazaki es menor en los organismos eucariotas (100 a 200 nucleótidos) que en los procariotas (1000 a 2000 nucleótidos) .
- Existen tres ADN polimerasas en los procariotas y cinco en los eucariotas.
- La replicación tiene un único origen en los procariotas, mientras que en los eucariotas existen múltiples (cientos en cada cromosoma de mamíferos, lo que hace que haya varios miles en el conjunto de su genoma). Cada unidad de replicación se denominareplicón y produce la síntesis de fragmentos de 100 a 150 nucleótidos. La necesidad de numerosos, puntos origen de la replicación resulta evidente, pues la cantidad de ADN en las células eucariotas es muchísimo mayor. Si sólo existiera un lugar de inicio, el proceso de replicación necesitaría varios meses para llevarse a cabo.
- La velocidad de replicación en cada replicón es menor en los eucariotas (hasta 50 veces) que en los procariotas.
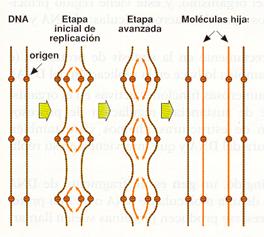
Replicación de los cromosomas (moléculas de ADN eucariótico) a partir de numerosos puntos iniciales.
3.1.4 Expresión de la información genética: transcripción y traducción. El código genético.
El ADN contiene información para que los aminoácidos se unan y formen las proteínas. Sin embargo, dado que la síntesis de proteínas se realiza en los ribosomas (situados en el citoplasma) y que el ADN se halla en el núcleo, del que no sale, se hace necesaria la existencia de alguna molécula que actúe como intermediario entre el ADN y los ribosomas. Este papel de intermediario lo realiza un tipo de ARN, el ARN mensajero (ARNm). El proceso de formación de los ARN se denomina transcripción.
Con la información contenida en la molécula de ARNm se puede sintetizar una cadena polipeptídica en un proceso denominado traducción que ocurre en los ribosomas. En este proceso intervienen otros tipos de ARN, el ARN ribosómico (ARNr), componente fundamental de los ribosomas, y el ARN de transferencia (ARNt), que transporta los aminoácidos hasta los ribosomas.
El flujo de información genética se puede expresar de la siguiente manera:
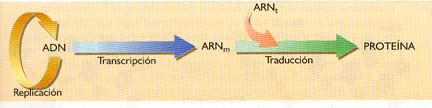
Este esquema fue considerado durante muchos años el "dogma central de la biología molecular", que postuló en 1970, Francis Crick.
Por supuesto, todo este trasvase de información se produce gracias a la naturaleza química de los ácidos nucleicos. Debido a la complementariedad de las bases nitrogenadas, el ADN se puede replicar y transcribir a ARNm, que se va a traducir por medio de los ARNt y ARNr, también gracias a la complementariedad de las bases.
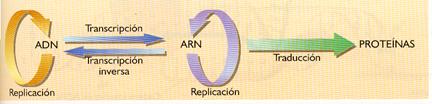
En la actualidad, esta forma de expresarlo ha tenido que ser modificada, debido a los mecanismos de replicación que presentan varios virus:
a) Algunos virus que almacenan su información genética en forma de ARN poseen una enzima, la ARN replicasa, capaz de fabricar copias de este ARN.
b) Los retrovirus almacenan su información genética en una molécula de ARN. Emplean una enzima, la transcriptasa inversa, que sintetiza ADN a partir de una molécula de ARN. El proceso recibe el nombre de retrotranscripción o transcripción inversa.
Tras el descubrimiento del comportamiento de estos virus, el dogma central de la biología molecular hubo de ser redefinido:
TRANSCRIPCIÓN
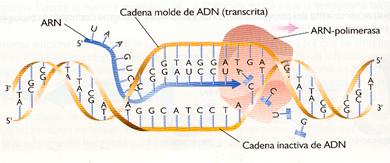
La síntesis del ARN o transcripción ocurre en el interior del núcleo. Como requisitos previos necesita:
a) Una cadena de ADN que actúe como molde . De las dos cadenas de nucleótidos que forman el gen, solo una, la denominada molde, se transcribe realmente, mientras que la otra, llamada informativa, no lo hace.
b) Enzimas . El proceso está catalizado por las ARN-polimerasas. En los procariontes solo existe una, mientras que en los eucariontes existen tres, llamadas ARN-polimerasas I, II y III: la I interviene en la formación del ARNr, la II lo hace en la síntesis de todos los ARNm y la III en la del ARNt y de un ARNr de pequeño tamaño.
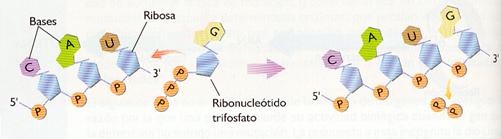
c) Ribonucleótidos trifosfato de A, G, C y U. Se unen mediante un enlace éster entre el ácido fosfórico situado en la posición 5' de un ribonucleótido trifosfato y el grupo -OH situado en posición 3' del último ribonucleótido de la cadena de ARN en formación.
El proceso de la transcripción
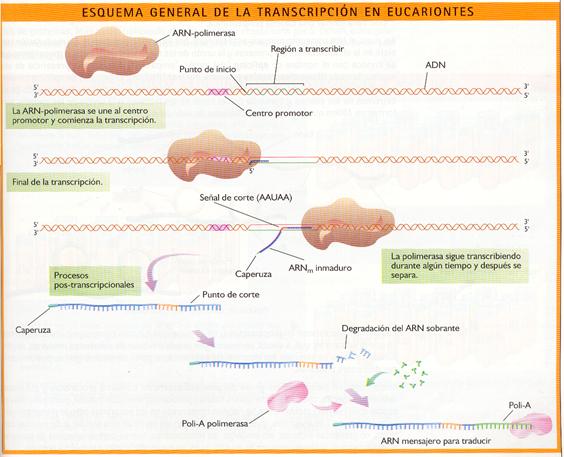
La transcripción consta de tres etapas: la iniciación, la elongación y la terminación. Tras ella se produce la maduración del ARN.
- Iniciación
Comienza cuando la ARN-polimerasa reconoce en el ADN que se va a transcribir una señal que indica el inicio del proceso. Tales señales, denominadas centros promotores, son unas determinadas secuencias cortas de bases nitrogenadas a las que se une la ARN-polimerasa.
La ARN-polimerasa hace que la doble hélice de ADN se abra para permitir que quede expuesta la secuencia de bases del ADN y se puedan incorporar los ribonucleótidos que se van a unir.
- Elongación
Es la adición de sucesivos ribonucleótidos para formar el ARN. La ARN-polimerasa avanza a lo largo de la cadena de ADN "leyéndola" en sentido 3'--5', mientras que el sentido de síntesis del ARN es 5'--3'. La enzima selecciona el ribonucleótido trifosfato cuya base es complementaria con la de la cadena de ADN que actúa como molde y lo une, mediante un enlace éster, al siguiente nucleótido, desprendiéndose un grupo pirofosfato (PPi).
En los eucariontes, tras la unión de los 30 primeros ribonucleótidos se añade en el extremo 5' una "caperuza" formada por metil-guanosín-fosfato, que durante la traducción será una señal de reconocimiento del inicio de lectura.
- Terminación
La ARN-polimerasa reconoce en el ADN unas señales determinación que indican el final de la transcripción. Esto implica el cierre de la burbuja formada en el ADN y la separación de la ARN-polimerasa del ARN transcrito.
- En los procariontes, la señal de terminación es una secuencia de bases palindrómica (secuencias que tienen la misma lectura de izquierda a derecha y de derecha a izquierda) formada por G y C seguidas de varias T, que origina al final del ARN un bucle. Este favorece su separación del ADN. El bucle se forma por autocomplementariedad de las bases G y C situadas en la cola del ARN.
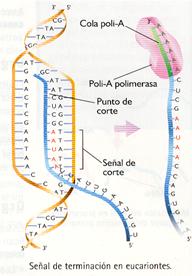
- En los eucariontes, la ARN-polimerasa transcribe regiones de ADN largas, que exceden la longitud de la secuencia que codifica la proteína. En ciertos puntos, una enzima corta el fragmento de ARN que lleva la información para sintetizar la proteína del ARN que sigue transcribiéndose. La señal de corte es una secuencia (AAUAA) que aparece sobre el ARN unos pocos nucleótidos antes del punto de corte, además de otras secuencias mal conocidas. Con posterioridad a la separación del ARN, una enzima (poli-A polimerasa) añade en el extremo final 3' una secuencia formada por unos 200 nucleótidos de adenina, llamada cola poli-A, que al parecer interviene en los procesos de maduración y transporte del ARN fuera del núcleo.
A veces, los ARNm no se pueden traducir directamente en proteínas, sino que necesitan un procesamiento previo o maduración postranscripcional.
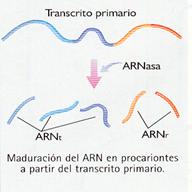
Organismos procariontes
El ARNm de los procariontes puede ser directamente traducido ya partir de él se forma una proteína funcional. No se puede hablar, por tanto, de una maduración de los mensajeros en estos organismos.
|
|
Sin embargo, cuando se transcribe el ADN que codifica los ARNt y los ARNr se forma una larga molécula de ARN que contiene numerosas copias de las secuencias del ARNr o el ARNt. Esta larga molécula, el transcrito primario, es posteriormente cortada en fragmentos más pequeños por enzimas específicas, para dar lugar a los distintos ARNt y ARNr. |
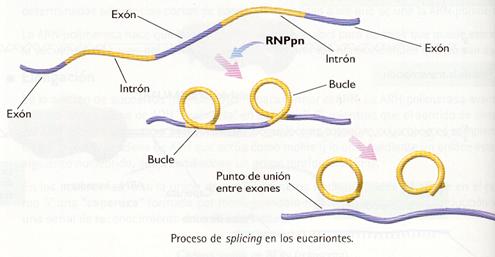
Organismos eucariontes
En los eucariontes la maduración es más compleja, ya que la mayor parte de los genes que codifican las proteínas están fragmentados. Cada gen consta de varios fragmentos denominados intrones y exones, intercalados unos con otros.
. Los intrones son secuencias de bases más o menos largas que se transcriben, pero que no se traducen, es decir, no codifican una secuencia de aminoácidos.
. Los exones son las secuencias que se transcriben y se traducen, es decir, tienen información para formar una cadena polipeptídica.
Así pues, el ARN transcrito primario está formado por intrones y por exones. Su maduración consiste en la eliminación de los primeros y la unión de los segundos mediante un mecanismo que se conoce con el nombre de splicing (del inglés, "empalme"). Requiere la presencia de una enzima llamada ribonucleoproteína pequeña nuclear (RNPpn). El proceso de splicing comienza cuando las secuencias intrónicas forman unos bucles que provocan el acercamiento de los extremos de los exones y continúa con el corte de los intrones y la unión de los exones, para formar un ARNm que ya está en condiciones de salir del núcleo.
Los intrones no existen en procariontes y no se sabe qué función cumplen en los eucariontes. Lo que sí se sabe es que, a veces, un mismo gen puede madurar de diferentes maneras, dependiendo de cómo se eliminen los intrones. De este modo, a partir de un solo gen se pueden obtener diferentes proteínas.
Actualmente se piensa que los genes del primitivo antecesor común a procariotas y eucariotas debían de tener intrones. Las bacterias los habrían perdido por selección natural, pues para ellas es crucial dividirse rápidamente. Se habrían conservado en las eucariotas porque presentan ventajas evolutivas. Las levaduras, que son eucariotas con un modo de vida similar al de muchas bacterias, no presentan intrones; sin embargo, las mitocondrias, que se cree descienden de bacterias endosimbiontes, sí tienen intrones en su ADN, pues no están sometidas a la misma presión.
EL CODIGO GENÉTICO
Una vez obtenida una copia del mensaje genético en forma de cadena de ARNm, ésta dirige la síntesis de proteínas en los ribosomas. Para ello, estos orgánulos interpretan la secuencia concreta de nucleótidos existente en la molécula de ARNm como la información necesaria para la unión de los aminoácidos precisos para constituir la proteína específica.
Consiste, pues, en una equivalencia entre dos polímeros específicos. Uno de ellos, el ARN, tiene dispuestas sus bases nitrogenadas en una secuencia concreta que contiene la información que determina el orden en que han de engancharse los sucesivos aminoácidos que forman la cadena polipeptídica. Por tanto, los ARNm con secuencias de bases nitrogenadas distintas llevan información para la síntesis de proteínas diferentes.
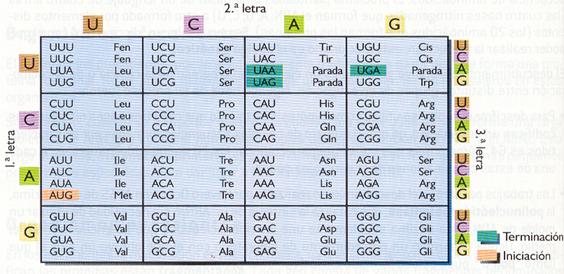
El código genético es, en definitiva, la clave que permite la traducción del mensaje genético a su forma funcional, las proteínas. Como sólo hay cuatro bases nitrogenadas distintas, las señales codificadoras para los 20 aminoácidos proteicos deben estar constituidas por más de una base. Si cada señal estuviera formada por dos bases nitrogenadas, sólo codificarían 42 = 16 aminoácidos, por la que aún quedarían aminoácidos sin codificar. Por tanto, cada señal que codifica para un aminoácido está constituida por tres bases nitrogenadas consecutivas (un triplete), es decir, 43 = 64 tripletes de bases distintas. George Gamow, creador de la teoría del big-bang sobre el origen del universo, fue el primero en formular este razonamiento teórico.
Los tripletes de bases del ARNm reciben el nombre de codones . Los tripletes del ADN correspondientes, que hallan sido transcritos, se denominan codógenos. Existen 61 codones codificadores de aminoácidos y 3 (UAA, UAG y UGA, llamados sin sentido) que señalan el final del mensaje y no especifican ningún aminoácido. Hay también un codón (AUG) que, además de codificar para el aminoácido metionina, es la señal de comienzo.
Este código genético presenta unas características que ayudan al cumplimiento de su función:
- Es universal. El código es compartido por todos los organismos conocidos, incluyendo los virus; así, por ejemplo, el codón UUG codifica para el aminoácido leucina tanto en los procariontes como en los eucariontes, lo mismo que ocurre con todos los codones. Este hecho indica que el código ha tenido un solo origen evolutivo.
Gracias a la genética molecular, recientemente se ha descubierto que esta universalidad tiene excepciones: concretamente, las mitocondrias y algunos protozoos, como Tetrahymena, utilizan un código genético ligeramente diferente.
- Es degenerado. Este término indica que la mayor parte de los aminoácidos, a excepción de la metionina y el triptófano, están codificados por más de un codón.
Los distintos codones que codifican para un mismo aminoácido se denominan codones sinónimos; esto supone una ventaja, ya que en el caso de que se produzcan cambios en algún nucleótido, es decir, que haya mutaciones, no se tiene por qué alterar el orden de los aminoácidos que forman una proteína.
- No presenta imperfección. Ningún codón codifica más de un aminoácido; lo contrario conllevaría problemas considerables, pues a partir de un gen se sintetizarían proteínas diferentes.
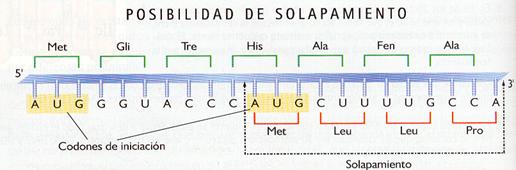
- Carece de solapamiento. Los tripletes de bases se hallan dispuestos de manera lineal y continua, sin que entre ellos existan comas ni espacios y sin que compartan ninguna base nitrogenada. Su lectura se hace en un solo sentido (5'®3'), desde el codón que indica el comienzo de la proteína hasta el que indica su final. Sin embargo, existe la posibilidad de que un mismo ARNm contenga varios codones de iniciación. Esto significaría que se podrían realizar varias fases de lectura y se sintetizaría más de un polipéptido.
TRADUCCIÓN O BIOSÍNTESIS DE PROTEINAS
La traducción es la transformación del mensaje de ARN en la síntesis de las proteínas correspondientes. Para que se produzca la traducción, se necesitan:
- ARNm.
- Los 20 aminoácidos.
- ARNt correspondientes a los 20 aminoácidos.
- Enzimas, factores proteicos y energía (GTP).
- Ribosomas. Los ribosomas son muy semejantes en organismos procarióticos y eucarióticos. Cada ribosoma tiene un sitio de unión al ARNm y tres sitios de unión al ARNt, el sitio A (aminoacil), el sitio P (peptidil) y el sitio E (salida=exit). Están formados por dos subunidades compuestas de moléculas de ARNr y muchas proteínas. La subunidad pequeña se une al ARNm y a los ARNt, y la subunidad grande es la encargada de formar el enlace peptídico entre los aminoácidos contiguos gracias a la enzima peptidil-sintetasa que contiene.
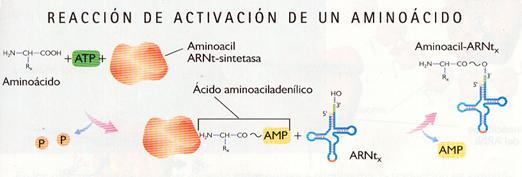
Activación de los aminoácidos
Los tripletes del ARN no reconocen directamente a los aminoácidos que especifican. La traducción del mensaje depende de la presencia de moléculas de ARNt que sean capaces de realizar ambos reconocimientos: en un extremo llevan unido el aminoácido, y el anticodón del brazo central es complementario al codón del aminoácido.
Cada aminoácido se activa por su aminoacil-ARNt-sintetasa correspondiente. Hay 20 enzimas distintas, una para cada aminoácido
Aminoácido + ATP + ARNt ——› aminoacil ARNt + AMP + PPi
Aminoacil-ARNt-sintetas
Esta reacción es imprescindible para la síntesis de proteínas
Fases de la síntesis proteica
La traducción o síntesis de proteínas tiene tres fases:
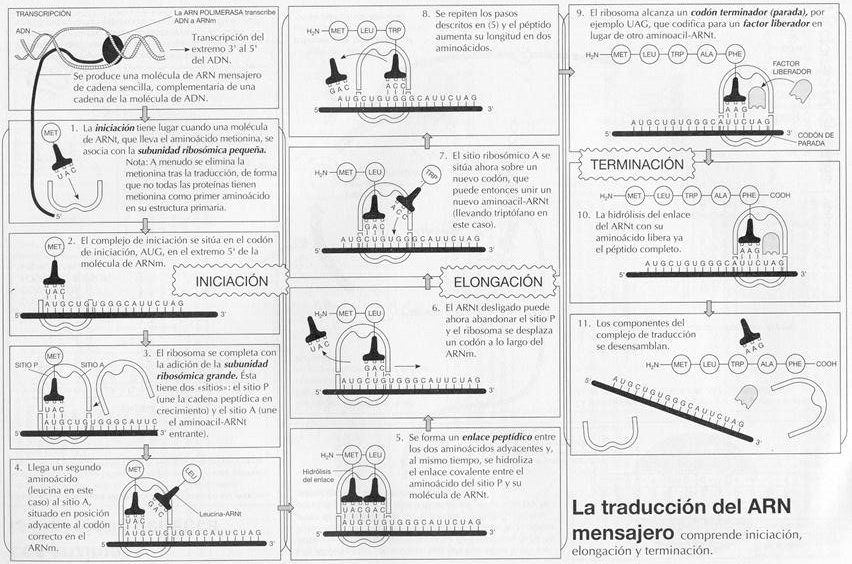
a) Iniciación : En esta fase la subunidad pequeña del ribosoma se une al extremo 5' del ARNm, colocándose el codón de iniciación (AUG) a la altura del sitio P, que es precisamente ocupado por el aminoacil-ARNt complementario (UAC), que porta el aminoácido Met (metionina) en eucariontes y el formilMet (formilmetionina) en procariontes (aminoácidos que normalmente se separarán una vez completada la síntesis del polipéptido). El conjunto de la subunidad ribosómica, el ARNm y el ARNt forma el complejo iniciador. A continuación, la subunidad mayor del ribosoma se une al complejo. Esta fase está catalizada por proteínas denominadas factores de iniciación (IF).
b) Elongación, que podemos dividir a su vez en tres subfases:
1. El siguiente codón del ARNm está expuesto a la altura del sitio A, lo que hace que allí también se coloque el aminoacil-ARNt correspondiente.
2. Una enzima, la peptidil transferasa, cataliza el mecanismo por el que el aminoácido situado en P se separa de su ARNt y se une con un enlace peptídico al aminoácido incorporado en A.
3. Al tiempo que el aminoacil-ARNt en sitio A (ahora con dos aminoácidos) es desplazado hacia el sitio P, de donde es expulsado el ARNt (ya sin aminoácido) que allí estaba, el ribosoma avanza exactamente tres nucleótidos a lo largo del ARNm, dejando un nuevo codón expuesto en el sitio A, permitiendo así que el ciclo comience de nuevo. Diferentes factores de elongación (EF) actúan como catalizadores en esta fase.
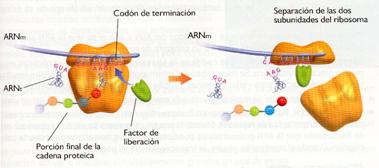
c) Terminación: Tiene lugar cuando en el sitio A se expone un codón de terminación para el que no existe ningún ARNt complementario. Lo que sí se sitúa en el sitio A va a ser un factor de separación (RF) que cataliza la separación del polipéptido. A continuación se separa el ARNm, así como las dos subunidades del ribosoma.
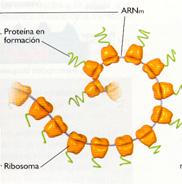
La síntesis completa de una proteína tarda entre veinte y sesenta segundos, pero lo más importante es que la traducción de un determinado ARNm implica la participación de numerosos ribosomas actuando simultáneamente, aunque en diferentes puntos del ARNm. Esto es, una vez que el primer ribosoma se aleja unos ochenta nucleótidos del codón de iniciación, un nuevo ribosoma inicia la traducción, y así sucesivamente, dando lugar a una estructura conocida como polirribosoma o, simplemente, polisoma, consistente en una hebra de ARNm unida a numerosos ribosomas como si de un collar se tratase.
Esquema de la traducción.
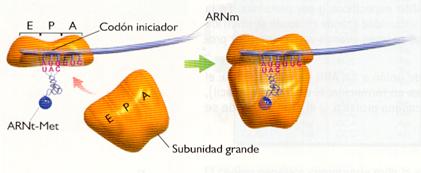
Iniciación.
|
Elongación. |
|
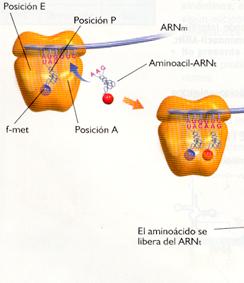
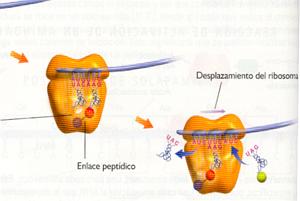
|
|
|
|
|
Terminación |
Polirribosoma |
Diferencias de la traducción entre procariontes y eucariontes
El proceso es prácticamente igual en ambos casos, pero presenta algunas diferencias dignas de mención:
- el primer aminoácido, como sé ha mencionado, es Met en eucariontes y formal-Met en procariontes;
- en eucariontes la transcripción y la traducción se llevan a cabo en espacios separados y, por tanto, de forma independiente, mientras que en procariontes la traducción puede comenzar cuando aún no haya acabado la transcripción;
- los diversos tipos de factores son diferentes.
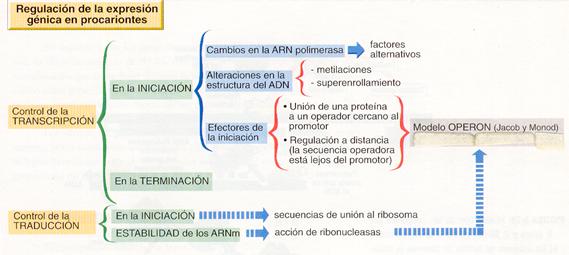
REGULACION DE LA EXPRESIÓN GENICA
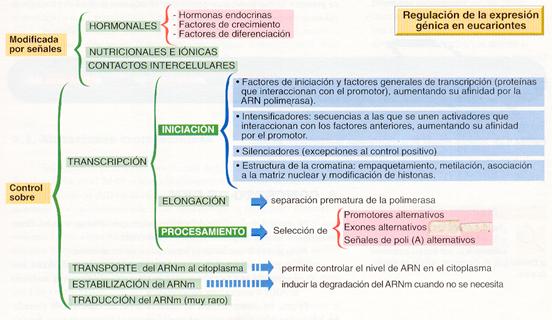
3.2. Alteraciones de la información genética.
El material genético no se mantiene inmutable generación tras generación; de forma inesperada y aleatoria, se producen alteraciones en el ADN (mutaciones) que pueden tener importantes consecuencias para el individuo en el que se manifiestan o pasar inadvertidas. Estas alteraciones, por otra parte, pueden ser negativas para el individuo, pero enormemente ventajosas para la especie a la que pertenece, ya que permiten aumentar la variabilidad genética, sin la cual no habría sido posible la evolución de los seres vivos por selección natural.
3.2.1. Concepto de mutación.
En 1901, Hugo de Vries creó el término mutación para referirse a los cambios repentinos aparecidos en individuos de la especie vegetal Oenothera lamarckiana con la que estaba trabajando. Sin embargo, el verdadero estudio genético de las mutaciones no comenzó hasta 1943, con las investigaciones de Salvador Luria y Max Delbrück, que contribuyeron de manera decisiva al esclarecimiento de sus causas y de su importancia biológica.
Las repercusiones biológicas de las mutaciones se derivan de la alteración que se produce en la secuencia de bases nitrogenadas del ADN, que se traduce, a su vez, en un cambio en la secuencia de los aminoácidos que constituyen la proteína correspondiente. De esta forma, la proteína codificada por ese ADN puede cambiar su función biológica o actuar incorrectamente. Si el segmento de ADN alterado corresponde a una zona reguladora, se puede modificar la expresión de otros genes.
Hoy se conoce perfectamente el papel que desempeñan las mutaciones en numerosos procesos biológicos, desde la evolución de las especies hasta el desarrollo del cáncer. Además, los individuos mutantes constituyen una valiosa fuente de información en multitud de trabajos de investigación sobre el material genérico.
Las mutaciones se pueden clasificar atendiendo a varios criterios que se recogen en el cuadro siguiente:
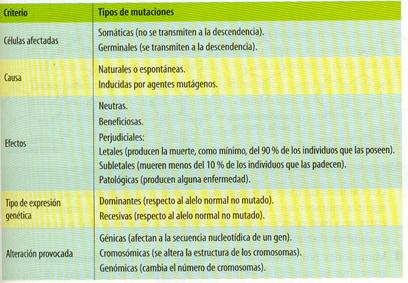
Según la extensión del material genético afectado, podemos clasificarlas mutaciones en:
MUTACIONES GENICAS
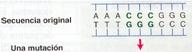
Las mutaciones génicas son las que alteran la secuencia de nucleótidos de un solo gen, por lo que se denominan puntuales. Se pueden distinguir dos tipos de mutaciones génicas: las sustituciones de bases y las mutaciones por corrimiento de la pauta de lectura.
- Sustituciones de bases
Suponen alrededor del 20% de las mutaciones génicas. Consisten en el cambio de una base del ADN por otra. Pueden ser:
- Transiciones. Si se sustituye una base púrica por otra púrica o bien una pirimidínica por otra pirimidínica.
- Transversiones. Si la sustitución es de una base púrica por otra pirimidínica, o viceversa.
Cualquiera de estas mutaciones afecta a uno solo de los nucleótidos y solo un triplete de bases es el que se ve afectado. Como el código genético es degenerado, el triplete puede sustituirse por otro que codifique al mismo aminoácido, de modo que la mutación no afectara al individuo y sería entonces una mutación silenciosa.
Por otra parte, puede que el nuevo triplete codifique otro aminoácido diferente. En este caso, salvo que sea un aminoácido que conforman el centro activo de una proteína, no tienen graves consecuencias. Si la mutación ocurre en el codón de terminación, se producirá una proteína más larga, hasta que aparezca un nuevo codón de terminación. Si la mutación crea un codón de terminación antes del lugar apropiado, se formará una proteína más corta.
En algún caso se puede producir una proteína que mejore a la original, y entonces el portador tendrá una ventaja que podrá transmitir a sus descendientes.
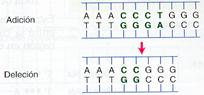
- Mutaciones por corrimiento de la pauta de lectura
Se denominan inserciones y deleciones si consisten en la adición o en la pérdida de algún nucleótido en la molécula de ADN, respectivamente. A partir del punto en el que inserción o deleción varían todos los tripletes de bases, por ello se dice que estas mutaciones provocan un corrimiento de la pauta de lectura. Cuando el gen afectado se traduce produce una proteína completamente diferente.
|
|
|

MUTACIONES CROMOSÓMICAS
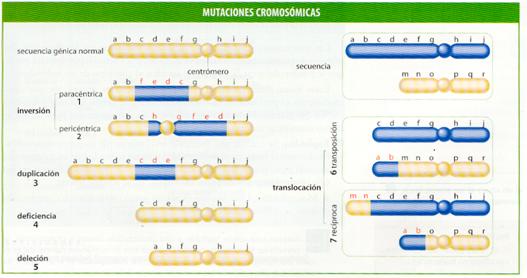
Este tipo de mutaciones afecta a la estructura de los cromosomas, por lo que es posible detectarlas al microscopio. La secuencia de bases nitrogenadas de los genes no está alterada, pero existen cambios en el número de genes o en su disposición lineal en los cromosomas. Se diferencian dos tipos de mutaciones según que la alteración afecte al número o al orden de los genes en los cromosomas:
· Alteraciones por la existencia de un número incorrecto de genes. Tienen lugar por un fallo en el apareamiento meiótico que puede producir un sobrecruzamiento erróneo, quedando un cromosoma con un fragmento extra y el otro con un déficit. Los gametos obtenidos originarán, tras la fecundación, diversas anomalías, como las siguientes:
- Deficiencias y deleciones . Consisten en la pérdida de un fragmento del cromosoma y, en consecuencia, de algunos genes, ya sea en el extremo (deficiencia) o en otro lugar (deleción).
- Duplicaciones . Un segmento de un cromosoma se encuentra repetido, por lo que existe un exceso de los genes correspondientes. En términos evolutivos, las duplicaciones poseen una importancia extraordinaria, ya que el aumento del número de genes puede determinar la aparición de nuevas variantes génicas en mutaciones posteriores.
· Alteraciones en el orden de los genes. Aunque estas mutaciones no ocasionan un perjuicio al individuo que las padece, producen gametos anormales que originarán una descendencia con déficit o exceso de genes. Se distinguen dos tipos:
- Inversiones . La disposición de los genes de un fragmento cromosómico está invertida. Si el fragmento invertido incluye el centrómero, la inversión se denomina pericéntrica, y en caso contrario, paracéntrica.
- Translocaciones . Un fragmento cromosómico cambia de posición, trasladándose a otro lugar del mismo cromosoma, a su homólogo o a otro cualquiera. Si la translocación se produce de un cromosoma a otro y de éste al primero se denomina recíproca; si el segmento simplemente pasa a situarse en otro cromosoma se llama transposición.
MUTACIONES GENÓMICAS O NUMÉRICAS
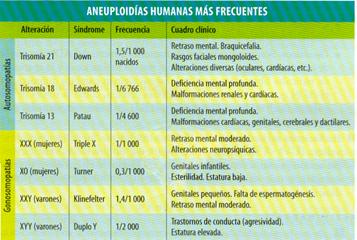
Las mutaciones genómicas o numéricas consisten en la alteración del número de cromosomas de una especie, ya sea por exceso o por defecto, por lo que se pueden detectar fácilmente al estudiar el cariotipo de un individuo.
Se distinguen dos tipos de mutaciones numéricas: euploidías y aneuploidías.
· Euploidías. Se trata de una alteración en el número de juegos cromosómicos. Se denomina juego cromosómico al conjunto formado por un cromosoma de cada tipo, por lo que los individuos diploides normales tienen en sus células somáticas dos de ellos. Las euploidías se subdividen en:
- Monoploidías . Únicamente existe un juego cromosómico completo, es decir, n cromosomas.
- Poliploidías . La anomalía consiste en la existencia de más de dos juegos cromosómicos. Las poliploidías pueden ser triploidías (con 3n cromosomas), tetraploidías (con 4n), hexaploidías (con 6n), etc.
Estas mutaciones ocasionan un aumento del tamaño celular. Los organismos poliploides pueden ser autopoliploides , cuando todas las dotaciones cromosómicas pertenecen a la misma especie, o
alopoliploides, cuando se han producido por hibridación de distintas especies.
·- Aneuploidías. No existe alteración del número de juegos cromosómicos completos. Solamente falta o sobra algún cromosoma. Las aneuploidías pueden ser:
- Nulisomías:(2n- 2) cromosomas. Falta una pareja cromosómica, por lo que esta alteración tiene efectos letales.
- Monosomías:(2n- 1) cromosomas. Falta un cromosoma de una determinada pareja.
- Trisomías: (2n + 1) cromosomas. Un cromosoma se encuentra por triplicado. Es frecuente en las plantas, donde provoca cambios morfológicos.
- Tetrasomías:(2n + 2) cromosomas. Existen cuatro ejemplares de un cromosoma determinado.
Las aneuploidías se producen por la fusión de un gameto normal (con n cromosomas) con otro que posee (n - 1), (n + 1) o (n + 2) cromosomas. Las más tolerables son las que afectan a cromosomas pequeños o a los cromosomas sexuales
3.2.2. Causas de las mutaciones.
Gran parte de las mutaciones se producen de manera espontánea, es decir, por causas naturales como errores que pueden ocurrir en la replicación o en la meiosis, o por cambios químicos espontáneos en el ADN.
La tasa de mutación espontánea, en general, es más baja en las bacterias y en otros microorganismos que en organismos más complejos. En la especie humana y en otros organismos pluricelulares se estima que ocurre una mutación en uno de cada cien mil a un millón de gametos, aunque existe una variación considerable de un gen a otro.
Otras mutaciones son causadas por la presencia en el medio de agentes físicos o químicos que pueden afectar a la estructura del ADN. Estas son mutaciones inducidas, y los agentes que las desencadenan son agentes mutagénicos.
AGENTES MUTAGÉNICOS
Desde que Hermann J. Muller ( 1927 ) y Lewis Stadler ( 1928) comprobaron que la aplicación de rayos X sobre las moscas de la fruta y sobre el centeno, respectivamente, inducía la aparición de mutaciones, se han descubierto muchos agentes mutagénicos, que se pueden clasificar en tres grupos: físicos, químicos y biológicos.
Agentes mutagénicos físicos
Aunque las subidas intensas y rápidas de la temperatura pueden provocar mutaciones, los agentes físicos mutagénicos por excelencia son las radiaciones, que se dividen en ionizantes y no ionizantes.
· Radiaciones ionizantes. Son radiaciones de longitud de onda muy corta y, por tanto, muy energéticas, que provocan la ionización de los átomos de las sustancias que atraviesan. Entre estas radiaciones se encuentran los rayos X y g, así como las partículas α y b y los neutrones emitidos en procesos radiactivos.
Los efectos de las radiaciones ionizantes sobre los seres vivos son de tres tipos:
- Fisiológicos . Pueden producir cambios enzimáticos que se traducen en modificaciones metabólicas.
- Citogenéticos . Comportan alteraciones en la estructura de los cromosomas, como deleciones y translocaciones.
- Genéticos . Son producidos por ionizaciones directas del ADN o a través de otros iones que, a su vez, provocan nuevas ionizaciones y la aparición de radicales libres muy reactivos. Éstos originan cambios químicos en el ADN que se traducen en mutaciones génicas, como la rotura de los enlaces nucleotídicos, la rotura y pérdida de bases nitrogenadas y la aparición de formas tautoméricas.
· Radiaciones no ionizantes. Son, fundamentalmente, las radiaciones ultravioleta (UV). A diferencia de las anteriores, no producen ionizaciones. Su acción primaria consiste en provocar el paso de electrones a niveles energéticos más altos, lo cual puede dar lugar a formas tautoméricas y dímeros de timina.
Agentes mutagénicos químicos
Numerosas sustancias tienen acción mutagénica (hidrocarburos policíclicos, aminas aromáticas, agentes alquilantes, colorantes industriales, pesticidas, etc.). A diferencia de las radiaciones, sus efectos, entre los que se pueden destacar los siguientes, suelen ser más retardados:
· Modificaciones de las bases nitrogenadas. Comprenden las reacciones de desaminación, alquilación e hidroxilación, que provocan emparejamientos erróneos. Así, por ejemplo, la desaminación de la adenina da lugar a la hipoxantina, que se aparea con la citosina en lugar de la timina; la guanina se transforma por alquilación en 6-O-metilguanina, que se aparea con timina en lugar de citosina.
· Sustituciones de bases. Están causadas por análogos debases nitrogenadas que provocan un emparejamiento erróneo durante la replicación al cambiar una base por otra. Entre ellos se encuentran la 2-aminopurina y el bromouracilo.
· Introducción de ciertas moléculas en la cadena polinucleotídica del ADN. Estas inserciones provocan la aparición de un exceso de nucleótidos en la hebra de nueva formación durante la replicación. De este modo, a partir de ese punto los tripletes de bases se alteran y el mensaje genético cambia.
Agentes mutagénicos biológicos
Algunos agentes biológicos aumentan la frecuencia de la mutación génica, Destacan entre ellos ciertos virus que pueden producir cambios en la expresión de algunos genes (por ejemplo, los retrovirus, los adenovirus o el virus de la hepatitis B humana, entre otros) y los transposones. Estos últimos son segmentos móviles de ADN que pueden cambiar de posición, trasladándose a otro lugar distinto dentro del mismo cromosoma o incluso a otro cromosoma.
Estos elementos móviles se han encontrado en todo tipo de organismos (maíz, levaduras, insectos, bacterias, etc.) y pueden originar mutaciones, ya que causan una activación o inactivación génica no deseada al insertarse en los genes estructurales o en los reguladores. Se cree además, que los virus mutagénicos podrían realizar su acción al llevar en su genoma transposones tomados de una célula previamente infectada que incorporarían a la nueva célula parasitada,
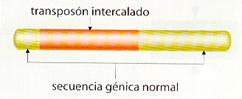
Transposon intercalado en una secuencia génica normal.
CAUSAS DE LAS MUTACIONES GÉNICAS
Si consideramos, que las mutaciones génicas son las mutaciones en sentido estricto, el estudio de las causas de estas, se puede considerar como el estudio de las causas de las mutaciones. Los otros tipos de mutaciones, se consideran más bien como alteraciones del material genético.
Las mutaciones génicas pueden producirse por tres causas: por errores de lectura durante la replicación del ADN, por lesiones fortuitas, como, por ejemplo, la rotura del enlace que une una base nitrogenada a la desoxirribosa, o por transposiciones (cambios de posición) de ciertos segmentos del gen.
· Errores de lectura. Los errores de lectura que pueden aparecer durante la replicación del ADN pueden deberse a dos causas: a los cambios tautoméricos y a los cambios de fase.
a) Los cambios tautoméricos . Cada base nitrogenada puede presentarse en dos formas diferentes denominadas formas tautoméricas o tautómeros, una es la normal y la otra la rara. Ambas formas están en equilibrio, y espontáneamente se pasa de una a la otra, lo que se denomina cambio tautomérico. Esto, si sucede durante la replicación, implica mutaciones, ya que cambia la base complementaria en la nueva hebra de ADN. Por ejemplo, la forma normal de la G se complementa con la C, mientras que la forma rara de G, es decir, su forma tautomérica, lo hace con la T.
b) Los cambios de fase. Son deslizamientos de la hebra que se está formando sobre la hebra molde, de forma que quedan bucles al volverse a emparejar. El crecimiento sigue y la diferencia queda fijada, originándose así la mutación.
· Lesiones fortuitas. Las lesiones fortuitas son alteraciones de la estructura de uno o de varios nucleótidos, que aparecen de forma natural. Las más frecuentes son:
a) Despurinización . Pérdida de purinas por rotura del enlace entre éstas y las desoxirribosas. Se producen a razón de unas 5.000 a 10.000 por día, en cada célula humana.
b) Desaminación . Pérdida de grupos amino en las bases nitrogenadas, que entonces se emparejan con una distinta de la normal. Se producen unas 100 por genoma y día.
c) Dímero de timina . Enlace entre dos timinas contiguas. Generalmente provocado por los rayos ultravioleta de la radiación solar.
· Transposiciones. Son cambios de lugar espontáneos de determinados segmentos de ADN, los denominados elementos genéticos transponibles. Éstos pueden ser menores que un gen (como las llamadas secuencias de inserción), un gen, o un grupo de genes (como los denominados transposones). Las transposiciones pueden producir mutaciones génicas si el elemento genético transpuesto se sitúa dentro de un gen, o mutaciones cromosómicas si pasa a un lugar donde no hay un gen, ya sea dentro del mismo cromosoma o incluso a otro cromosoma.
3.2.3. Consecuencias de las mutaciones.
En los seres unicelulares, las mutaciones afectan al organismo, se heredan e incrementan la variación en las poblaciones. En los pluricelulares, las mutaciones tienen consecuencias distintas si se producen en las células germinales o si afectan a las células somáticas. En el primer caso, los efectos son heredables; pero en el segundo, que constituye las llamadas mutaciones somáticas, no lo son, puesto que los cambios sólo se producen en células que mueren con el individuo y, por lo tanto, no se transmiten a la descendencia. Estos hechos tienen importantes consecuencias biológicas.
3.2.3.1. Consecuencias evolutivas.
Las distintas teorías evolucionistas como explicaciones que son del origen de las especies, consideran fundamental el papel de los cambios genéticos, fuente de la variabilidad sobre la que actúan los mecanismos que regulan la evolución, el más aceptado de los cuales es la selección natural. Cuanto mayor sea el número de variantes, mayor número de posibilidades de evolución se ofrece a una especie o población.
Sin embargo, cada una de las diferentes teorías evolucionistas ofrece su particular punto de vista sobre el papel de las mutaciones en la evolución.
· Para los neodarwinistas, la mutación es una fuente de variación que proporciona beneficios o perjuicios. La selección natural elimina las mutaciones perjudiciales y favorece que las frecuencias de los genes beneficiosos se incrementen notablemente en la población, hasta producir un cambio en el tiempo que supone la consolidación de nuevas características; es decir, que implica una evolución.
· Para los neutralistas, la mayoría de las mutaciones no suponen ni perjuicio ni beneficio. Según ellos, estas determinan características neutras con respecto a la selección, y es el azar quien dirige, en gran medida, el proceso evolutivo.
· Por su parte, los partidarios de la teoría de los equilibrios interrumpidos (puntuados) consideran que existen mutaciones génicas pequeñas, que proporcionan la posibilidad de adaptaciones; y grandes cambios (cromosómicos, por ejemplo) que explican la aparición de características propias de los grandes grupos taxonómicos, como familias, clases, etc.
Sea como fuere, la mutación en sentido estricto es la primera fuente de variación, pero no la única. La recombinación genética incrementa el número de gametos distintos durante su formación en la meiosis, lo que produce mayores variantes en un proceso (la reproducción sexual) cuyo sentido y función es precisamente la aparición de variabilidad.
La importancia evolutiva de la recombinación y de los elementos genéticostransponibles puede reconocerse también en las bacterias. Los fenómenos parasexuales, como la conjugación, y la aparición de cepas que presentan resistencia a numerosos antibióticos, gracias a la transposición, son una forma de incrementar la velocidad de la evolución. Mutación, recombinación y transposición son las fuentes de la variación y, por lo tanto, el primer paso para la aparición de nuevas formas de vida.
También, las mutaciones somáticas pueden afectar al desarrollo de las capacidades que posee un individuo para mantenerse adaptado a su medio; y, aunque estas capacidades influyen en su supervivencia, no modifican la composición genética de la población.
La creciente aceptación de la relación existente entre genes y cáncer supone una importante vía para entender el origen de una de las enfermedades que más afectan a la humanidad.
Se ha demostrado que muchos tipos de cáncer son producidos por agentes ambientales que provocan mutaciones. Pero estas mutaciones son, en la mayoría de los casos, somáticas, con lo que se puede establecer una conclusión que sirve incluso para aclarar dos conceptos básicos en genética: el cáncer puede ser una enfermedad genética, pero, salvo en contadas ocasiones que constituyen una excepción, no es una enfermedad hereditaria.
3.2.3.2. Efectos perjudiciales.
Como acabamos de ver con el cáncer, las mutaciones pueden ser perjudiciales, y estos perjuicios se pueden estudiar dependiendo cuando se produce la mutación, ésta puede presentarse en las células somáticas, siendo el perjuicio para el individuo en el que se produce apareciendo enfermedades como el cáncer, que aunque pueden tener un determinante genético puede verse favorecido por agentes mutagénicos, ejemplo de esto lo podemos ver el cáncer de piel que producen las radiaciones solares.
Si la mutación afecta a las células germinales (gametos) los efectos se transmitirán a la descendencia, provocando la mayoría de las veces enfermedades, incluso abortos por causa de los llamados genes letales.
Las principales enfermedades producidas por mutaciones génicas son la anemia falciforme, la hemofilia, la acondroplasia (una de las causas del enanismo), etc.
Entre las mutaciones cromosómicas y genómicas, que ya dijimos que más bien se pueden considera como alteraciones, nos encontramos con enfermedades que se suelen llamar síndromes, así tenemos el síndrome de “Cri-du-chat”, que es una alteración que se produce por una deleción en el brazo corto del cromosoma 5, y su cuadro clínico es un llanto característico, cráneo pequeño, oligofrenia y retraso psicomotor. Los síndromes de Down, Turner, Klinefelter, etc. que son genómicas.
3.3. Genética aplicada.
La genética podemos utilizarla para la mejora de las especies animales y vegetales, siendo la ingeniería genética. La que más utilización se está haciendo hoy día.
Es bueno aplicar los conocimientos a favor de la humanidad, pero sin olvidarnos de la ética y del uso que hagamos no esté en contra de la naturaleza y la dignidad humana.
A continuación estudiaremos algunas de estas aplicaciones, que bien orientadas podrán curar enfermedades y paliar el hambre en el mundo con la mejora genética de especies importantísimas en la alimentación.
3.3.1. Enfermedades hereditarias: concepto.
Los primeros estudios en genética humana se realizaron investigando las causas de algunas enfermedades hereditarias, ya que una de las finalidades de la investigación genética es conseguir prevenirlas o paliar sus efectos en el ser humano.
Muchas veces se confunden los conceptos genético y hereditario. El término genético hace referencia a su relación con los genes, mientras que el término hereditario se refiere a la posibilidad de ser transmitido de una generación a la siguiente. Así, no todas las enfermedades genéticas son hereditarias.
Cuando el genoma de las células somáticas de un individuo sufre cualquier tipo de alteración, puede causarle enfermedades graves en algunas ocasiones, pero no serán heredadas por sus descendientes. Por ejemplo, una exposición prolongada a la radiación ultravioleta del Sol produce la alteración de determinados genes que causan algunos tipos de cáncer de piel. Aunque esta enfermedad se debe a una alteración genética, no es hereditaria.
Sin embargo, si la alteración genética ha ocurrido en las células germinales, sí se producirán enfermedades hereditarias, es decir, enfermedades que serán transmitidas de padres a hijos.
Otro concepto que suele confundirse con los anteriores es el de enfermedades congénitas, que son aquellas que se manifiestan desde el nacimiento de un individuo. Una enfermedad puede ser hereditaria y no ser congénita, es decir, no aparecer en el momento del nacimiento, sino en la edad adulta. Por ejemplo, el «Baile de San Vito» o «Corea de Huntington» es una enfermedad hereditaria que suele aparecer en la edad adulta; se manifiesta como una serie de contracciones descontroladas de los músculos que van haciéndose cada vez más severas y que conducen finalmente a la muerte del individuo.
Enfermedades ligadas alos autosomas
Cuando el gen causante de una enfermedad se encuentra en un cromosoma autosómico, su herencia tan solo depende de si está regulada por un alelo dominante o recesivo.
Alelos dominantes. Se manifiesta la enfermedad tanto en los homocigóticos como en los a heterocigóticos, por lo que siempre, que un individuo padezca la enfermedad, al menos uno de sus progenitores estará afectado por ella.
La sindactilia (dedos unidos por una membrana), la polidactilia (poseer más de 5 dedos en cada extremidad) o la braquidactilia (dedos cortos) se deben a genes, dominantes localizados en autosomas.
Alelos recesivos. La enfermedad solo se manifiesta al encontrarse en estado homocigótico. Debido a esto pueden pasar muchas generaciones sin que aparezcan individuos que manifiesten la enfermedad correspondiente.
Es el caso de enfermedades tales como: albinismo (incapacidad para formar melanina, pigmento que colorea nuestra piel, pelo y ojos), sordomudez congénita, fibrosis quística y anemia falciforme.
Existen otras enfermedades hereditarias debidas a alteraciones pequeñísimas en la secuencia de un solo gen, que se expresa como un déficit de una proteína enzimática que intervendría en un determinado proceso y que consecuentemente origina una serie de alteraciones del proceso que se conocen como «errores innatos del metabolismo» o incapacidades del individuo afectado para realizar determinados procesos bioquímicos, siendo también hereditarios: la alcaptonuria, la fenilcetonuria, la enfermedad de Tay-Sachs y otros muchos son de este tipo.
Enfermedades ligadas a los cromosomas sexuales
Algunas enfermedades están producidas por genes que se encuentran en los cromosomas sexuales, La mayoría de ellas son debidas a alelos recesivos. En estos casos, la herencia dependerá del sexo del progenitor que lleve el gen que causa la enfermedad y del sexo de la descendencia.
· Genes localizados en el cromosoma X. La mujer podrá ser heterocigótica para los alelos de estos genes (portadora de la enfermedad) u homocigótica, y entonces manifestará la enfermedad. El hombre únicamente podrá ser homocigótico, ya que solo lleva un alelo para ese gen en su único cromosoma X. La hemofilia (incapacidad en mayor o menor grado para la coagulación sanguínea) y el daltonismo (incapacidad para distinguir los colores complementarios como el rojo y el verde) responden a este tipo de herencia. En cualquiera de estos casos, la posibilidad de que exista una mujer con la enfermedad es mucho menor que la estimada para un varón, ya que el alelo recesivo ha de encontrarse en los dos cromosomas X.
· Genes localizados en el cromosoma Y. Los caracteres localizados en el segmento diferencial del cromosoma y solo se presentarán en los hombres y se transmitirán a todos los hijos varones. Una enfermedad de este tipo es la ictiosis , que consiste en la formación de una piel muy gruesa y con escamas.
3.3.2. Mejora genética de animales y plantas.
El nacimiento de la genética estuvo ligado a la actividad de los agricultores y ganaderos que trataban de mejorar el rendimiento de las plantas de cultivo y animales domésticos. Los avances de la genética tienen una aplicación directa en la agricultura y la ganadería.
Objetivo de la mejora genética
La mejora genética pretende obtener los genotipos más adecuados para unas características ambientales determinadas y para el uso al que van a ser destinados. Esto significa:
- Aumentar el rendimiento o producción de animales de granja y plantas de cultivo.
- Mejorar la calidad de los productos obtenidos.
- Aumentar su resistencia ante plagas y enfermedades.
- Lograr su adaptación a otros ambientes y extender su área de producción.
Tradicionalmente, se ha utilizado la selección artificial como método para lograr estos objetivos. Se eligen deliberadamente a los individuos con las características deseadas y se utilizan como progenitores de la siguiente generación. No obstante, si el carácter que se quiere seleccionar está muy influido por el ambiente, la selección basada únicamente en el fenotipo puede no dar los resultados esperados.
3.3.2.1. Procedimientos clásicos.
La aplicación de las bases teóricas de la genética a la mejora de animales y plantas ha ampliado considerablemente las expectativas. Sin embargo, los resultados han sido más espectaculares en la mejora vegetal que en la animal a consecuencia de los problemas asociados a la cría de los animales (número limitado de descendientes, lento crecimiento, imposibilidad de autofecundación, costosa manipulación, etc.). La aplicación de técnicas más modernas, como la congelación de semen, inseminación artificial, ovulación múltiple, congelación de embriones o la fecundación in vitro, ha logrado paliar estos problemas.
· Hibridación y posterior reorganización génica. La hibridación es un método que resulta de la aplicación directa de la genética mendeliana. Consiste en:
-Seleccionar razas puras con los caracteres ventajosos que se desean encontrar juntos en el mismo individuo.
- Hibridar dos razas puras diferentes con los dos caracteres de interés.
- Permitir la autofecundación de la F1. Entre la gran variedad de descendientes, algunos presentarán juntos los caracteres ventajosos. Varias generaciones de autofecundación producirán homocigotos para los genes de interés.
· Inducción de mutaciones. El método anterior (hibridación) utiliza la variabilidad natural para lograr combinaciones más ventajosas. Sin embargo, se puede recurrir al uso de agentes mutagénicos para disponer de una mayor variabilidad.
La inducción de mutaciones se utiliza fundamentalmente en la mejora vegetal, y el procedimiento consiste en irradiar los granos de polen o las semillas. Las mutaciones dominantes aparecen en la primera generación, y las recesivas, tras sucesivas generaciones de autofecundación.
· Obtención artificial de poliploides. La poliploidía de los vegetales puede provocarse en el laboratorio mediante la utilización de la colchicina, sustancia que impide la formación del huso acromático en la división celular. Con la producción de poliploides se pretende fundamentalmente el aumento de tamaño y, con él, un mayor aprovechamiento económico. Con este método se obtienen:
- Autopoliploides , que tienen repetido varias veces el número de dotaciones cromosómicas. Por ejemplo, el centeno silvestre presenta una dotación cromosómica diploide (2n = 14) y la variedad tetraploide supera en calidad y producción a los diploides originales.
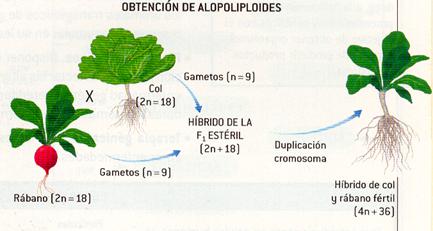
- Alopoliploides , que son híbridos (interespecíficos) de dos especies próximas, en los que la poliploidía provocada permite evitar la esterilidad. Un ejemplo es el híbrido de col y rábano. Fue todo un éxito como experimento genético, pero un fracaso total como experimento comercial.
3.3.2.2. Ingeniería genética.
La introducción de un gen foráneo en un organismo puede tener como finalidad la mejora. Los organismos eucarióticos modificados por ingeniería genética se conocen como organismos transgénicos.
- Animales transgénicos: Algunos caracteres de los animales domésticos pueden mejorar por ingeniería genética. Por ejemplo, se puede acelerar su crecimiento, mejorar la calidad de sus productos o aumentar su resistencia a las enfermedades.
- Plantas transgénicas: Las plantas de cultivo se están modificando actualmente para conseguir resistencia a la sequía, las heladas, insecticidas y herbicidas, o para la mejora de cualidades deseadas por los consumidores.
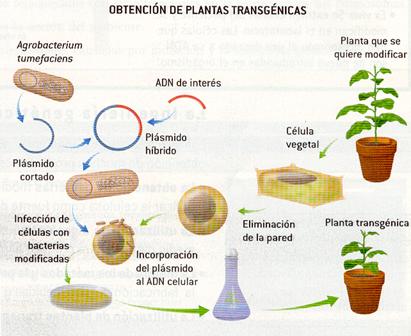
En las plantas se suele utilizar como vector un plásmido, inductor de tumores, de una bacteria (Agrobacterium tumefaciens). Esta bacteria infecta de forma natural a la planta y produce agallas o tumores que están formados por células transformadas por el plásmido.