







2.5.5.4.2.3. Respiración: ciclo de Krebs, cadena respiratoria y fosforilación oxidativa.
La respiración es el conjunto de procesos catabólicos que consisten en la degradación de sustancias orgánicas -combustible- para formar la energía que le célula necesita. Estas sustancias orgánicas pueden, ser de origen extracelular o sustancias de reserva almacenadas por la célula.
La obtención de energía puede realizarse con necesidad de oxigeno (respiración aerobia u oxibiótica), o en ausencia de dicha molécula (respiración anaerobia o anoxibiótica).
En la respiración aerobia el oxígeno molecular (O2) es el que acepta los hidrógenos para formar agua, y en la respiración anaerobia la sustancia que se reduce es diferente del oxígeno, pero en todo caso una sustancia inorgánica como el NO3-, el S04=, etc.
La energía desprendida en la respiración esta cuantificada, es decir, se desprende poco a poco en forma de moléculas de ATP, para que la célula pueda utilizarla y no le sea perjudicial.
La respiración aerobia, que tiene lugar en el citosol y en las mitocondrias, al ser degradados los combustibles de forma total (los productos finales son CO2 y H2O) el rendimiento energético es el máximo. Este tipo de respiración está generalizado a, prácticamente, todos los seres vivos.
La respiración anaerobia es mucho más ineficaz energéticamente que la anterior, pues los productos que quedan, contienen todavía energía. La presentan solamente algunas bacterias.
La respiración aerobia, la más importante, por ser la más empleada por los seres vivos, y la que más energía produce, se realiza en varias fases, que son: el ciclo de Krebs, la cadena respiratoria y la fosforilación oxidativa.
CICLO DE KREBS
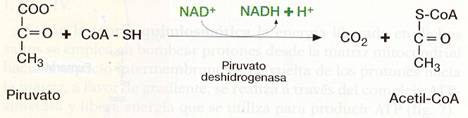
El piruvato obtenido en la glucólisis pasa, por transporte facilitado, a la matriz mitocondrial, donde se convierte en acetil-CoA mediante un proceso de oxidación y descarboxilación, en el que interviene el complejo multienzmático deshidrogenasa.
El grupo carboxilo se desprende formando CO2, y queda un grupo acetilo de carbonos que se une al a C o A y se oxida, al tiempo que el NAD+ se reduce a NADH + H+.
Esta reacción es irreversible y dirige al piruvato hacia su oxidación en el ciclo del Krebs.
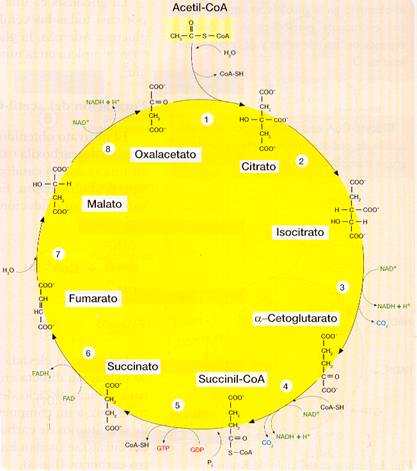
Este ciclo, consiste en una secuencia de reacciones a través de las cuales se lleva a cabo la oxidación final y total de la mayoría de los combustibles metabólicos. Se inicia con la incorporación al ciclo del acetil-CoA procedente de la glucosa (o, en otros casos, de ácidos grasos o aminoácidos). También se denomina ciclo del ácido cítrico o de los ácidos tricarboxílicos porque interviene el ácido cítrico (citrato en su forma aniónica) que posee tres grupos carboxilo (-COOH).
Este ciclo se considera el centro del metabolismo aerobio, en el que confluyen la mayoría de los procesos catabólicos e, incluso, algunas vías anabólicas (por eso se dice que es una ruta anfibólica). Tiene lugar en la matriz mitocondrial, donde se encuentran las enzimas necesarias para cada paso.
El acetil-CoA se incorpora al ciclo de Krebs y se producen 8 reacciones:
1. El grupo acetilo del acetil-CoA se condensa con el oxalacetato formando citrato, una molécula de 3 grupos carboxilos y 6 carbonos.
2. El citrato se isomeriza a isocitrato que es más fácilmente oxidable.
3. Por descarboxilación oxidativa el isocitrato se convierte en α-cetoglutarato. En esta reacción se desprende CO2 y se forma NADH + H+.
4. De nuevo tiene lugar una descarboxilación oxidativa en la que el α-cetoglutarato pasa a succinil-CoA. También aquí se desprende CO2 y se forma NADH + H+.
5. El succinil-CoA pierde la CoA y pasa a succinato, liberando la energía suficiente como para que se forme GTP mediante un proceso de fosforilación a partir del sustrato.
6. El succinato se oxida a fumarato, mediante una reacción acoplada en la que la coenzima FAD se reduce a FADH2.
7. Por hidratación del doble enlace del fumarato se forma malato.
8. A partir del malato se regenera el oxalacetato con el que se iniciaba el ciclo. La oxidación del malato se acopla a la formación de otra molécula de NADH + H+.
En resumen, la reacción global del ciclo de Krebs es la siguiente:
Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + H2O à 2 CO2 + CoA-SH + 3 (NADH + H+) + FADH2 + GTP
En cada vuelta del ciclo:
- Entra un grupo acetilo (dos átomos de carbono) que es oxidado completamente (por lo que salen del ciclo otros dos carbonos en forma de CO2).
- Tres moléculas de NAD+ son reducidas a NADH + H+.
- Una molécula de FAD es reducida a FADH2.
- Se forma una molécula de GTP equivalente al ATP.
A continuación, el NADH + H+ y el FADH2 se oxidan mediante la cadena de transporte electrónico mitocondrial generando ATP.
Esquema del ciclo de Krebs.
CADENA RESPIRATORIA Y FOSFORILACION OXIDATIVA
Aunque el ciclo de Krebs es característico de la respiración aerobia, habrás observado que en ninguna de sus reacciones interviene el oxígeno molecular. La utilización del oxígeno es exclusiva de esta última fase, en la que los electrones captados por el NADH+ H+ y el FADH2 son transportados hasta el oxígeno molecular para formar agua.
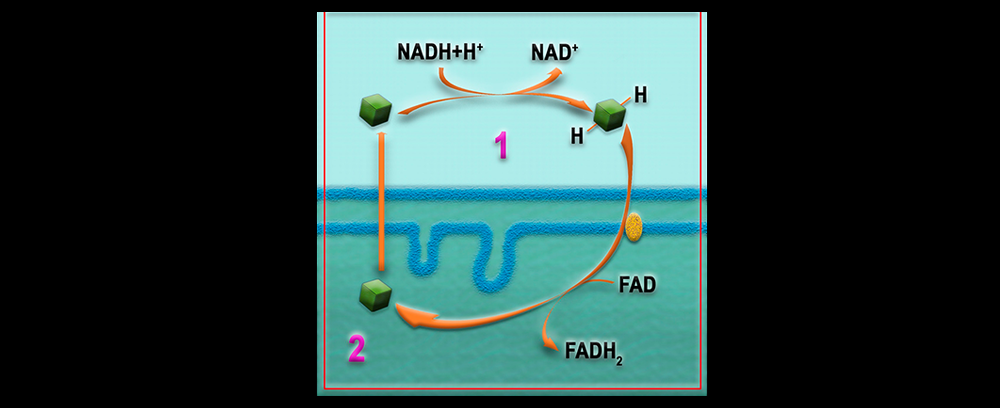
El transporte de electrones se inicia cuando una molécula de NADH + H+ o de FADH2 se oxida y cede H + y electrones a una de las, aproximadamente, 15 moléculas que forman la cadena de transporte electrónico situada en la membrana mitocondrial interna.
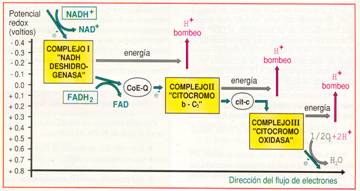
El transporte se realiza a través de una serie de reacciones de óxido-reducción. Cada reacción hace intervenir a dos constituyentes de la cadena respiratoria con potenciales redox diferentes. El potencial redox mide la afinidad de un transportador por los electrones. La afinidad es mayor cuanto mayor es el potencial redox. Los electrones se desplazan desde el constituyente que tiene potencial redox menor, hacia el que lo presenta mayor.
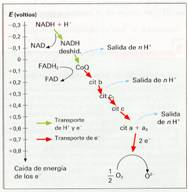
Las proteínas transportadoras están agrupadas en tres grandes complejos. Cada grupo posee un potencial redox más positivo que el anterior; los electrones descienden en cascada desde el NADH + H+ hacia los grandes complejos de enzimas, situados en niveles energéticos cada vez menores, hasta que finalmente son transferidos al oxígeno.
El NADH + H+ cede H+ y electrones al complejo I, mientras que el FADH2 los cede directamente al coenzima Q, al que también van a parar los del complejo I. El coenzima Q transfiere los electrones al complejo II que, a través del citocromo-c, pasan hasta el complejo III. Finalmente, los electrones reducen el oxígeno y forman agua.
|
|
|
|
Distintas representaciones de la cadena respiratoria. |
|
Entre las moléculas de la cadena respiratoria hay transportadores de electrones y transportadores de hidrógeno. Cuando un transportador de hidrógeno es oxidado por un transportador de electrones, los protones quedan libres en la matriz. La disposición ordenada y fija de los transportadores en la membrana permite utilizar la energía liberada en el transporte de electrones para bombear estos protones desde la matriz hacia el espacio intermembrana.
Hay tres puntos en la cadena respiratoria en los que ocurre esta translocación de protones que, debido a la impermeabilidad de la membrana mitocondrial interna, se acumulan en el espacio intermembrana. Así se origina un gradiente electroquímico de protones, es decir, un gradiente de concentración (o lo que es lo mismo, de pH) y un gradiente eléctrico (potencial de membrana), al ser diferente la distribución de H+ a ambos lados de la membrana.
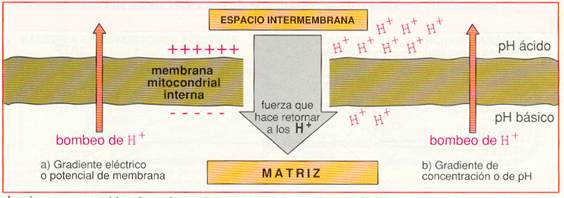
Los dos componentes del gradiente electroquímico se suman para impulsar a los H+ de vuelta a la matriz.
Debido a la impermeabilidad de la membrana interna, el retorno de protones a la matriz sólo puede hacerse a través de la ATP-sintetasa. Esta proteína utiliza la energía acumulada en el gradiente de H+ para fosforilar el ADP y transformarlo en ATP .La hipótesis que explica el acoplamiento de estos dos procesos, uno químico, de oxidación-reducción en la cadena respiratoria, y uno osmótico, de transporte de protones, se conoce con el nombre de hipótesis quimiosmótica y su resultado es la fosforilación oxidativa.
|
|
|
Transporte de electrones, translocación de protones y fosforilación a partir del NADH. |
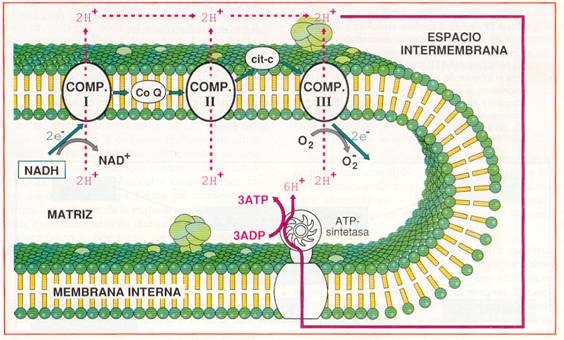
Cada NADH que llega a la cadena respiratoria cede una pareja de electrones, que en su transporte liberan energía suficiente para bombear 6 H+ desde la matriz al espacio intermembrana. Si los electrones proceden del FADH2 sólo se bombean 4 H+.
Por cada 2 H+ que vuelven a la matriz a través de la ATP-sintetasa, se fosforila un ADP. Por tanto, en la cadena respiratoria, se pueden obtener 3 ATP por cada NADH + H+ obtenido en el catabolismo y 2 ATP por cada FADH2.